植物の茎は、重力と反対方向の空に向かって伸び、根は重力にしたがって地中に伸びる。重力刺激に応じて、茎や根が成長する方向を変える運動は重力屈性と呼ばれ、進化論で有名なダーウィンの著書にも取り上げられている。重力屈性が関与する枝や根の成長角度は、農業生産量に直結する重要な形質でもあるが、植物が重力の方向を感知するしくみには多くの謎が残されている。変異体の探索と遺伝子解析から重力屈性のしくみを解き明かそうとしている森田美代教授に研究の背景と魅力を聞いた。
両親との賭け
高校の頃、夏休みも毎日部活動で学校に行っていた私は、生物の先生に頼まれて植物の水やりをすることになりました。それが縁でいろいろな植物の話を聞く機会を得たのですが、とても楽しそうに植物の話をする先生の影響で、生物学科への進学を希望するようになりました。しかし、娘の大学進学は想定外だった両親から課された条件は「家から通える公立大」。
瀬戸内海を超えて進学する周りの友達を横目で見ながら鬱々とする私に、生物の先生は「本当に勉強がしたいなら大学院という場所もある」と、それまで知らなかった大学院という存在を教えてくれました。そこで、私は両親にある“賭け”を持ちかけました。「もし私が4年後も生物学の勉強を続けたい気持ちを持ち続けていたら、大学院の修士課程に進学させてほしい。その時は自分で大学を選びたい。」
大学の授業でワトソンの教科書Molecular Biology of the Gene に出会い、分子生物学に興味を持つようになりました。卒業研究のテーマは出芽酵母の重金属耐性でした。酵母の銅耐性株では、重金属を結合して無害化する金属結合タンパク質の発現が銅によって誘導されます。環境の変化を細胞が認識して、それに対応して必要な遺伝子の転写を行う、正に教科書で学んだ転写調節を目の当たりにしたのです。その精緻な仕組みをとても合理的に説明できる分子生物学に美しさを感じ、より深く学びたいと思うようになっていた頃、指導教授から大学院進学を勧められました。「後輩が京大理学部で教授をしているけど、植物の分子生物学を始めるらしいよ。」その言葉だけで京都大学の理学研究科植物学専攻の受験を決めました。合格後、案の定両親と一悶着ありましたが、ともあれ瀬戸内海を越えることに成功しました。
植物学専攻のはずが・・・
「森田さんにはバクテリオファージの粒子形成の研究をしてもらうから」という教授の一言で、大学院での研究テーマが決まりました。後で分かったのですが、教授は、新しく迎える助教授と共に植物の分子生物学研究を始めるつもりだったようです。私の入学の翌年に助教授(田坂昌生先生(現・奈良先端科学技術大学院大学名誉教授))が着任し、同じ研究室でシロイヌナズナを用いた研究がスタートしました。私はシロイヌナズナの研究を横目で見ながら、ワトソンも研究したバクテリオファージ研究に邁進しました。一般に生物学研究では、先行研究を基に作業仮説を立てて実験を行い、その結果について考察し、さらに次の作業仮説を立ててという一連のサイクルを何度も積み重ね、それらを論理立てたストーリーとしてまとめて論文として発表します。一晩で増えるバクテリオファージはこのサイクルがとても早いものでした。大学院生というキャリアの早い時期に、このような研究の訓練を数多く行うことができたことは、とても得がたい経験になりました。
重力屈性との出会い
学位取得後、植物の研究に携わりたいと思いつつ、しばらく半官半民の会社で研究員として働きました。大腸菌を使って、mRNAの二次構造そのものがタンパク質を合成する効率を制御する熱センサーとして機能することを初めて実験的に証明したのですが、幸運なことに当時奈良先端大の教授になっておられた田坂先生にこの研究を評価していただき、「助手のポジションがあるのでシロイヌナズナの研究をしてみないか」とお声がけいただきました。32歳にしてようやく植物分野での研究がスタートしました。田坂研究室では、植物の形態形成と重力屈性の二本柱で研究を進めており、私は重力屈性の研究に携わることになりました。進化論で有名なダーウィンが研究したことでも知られている「屈性」は、光や重力、水分、接触などの環境刺激に応じて、茎や根が成長する方向を変えるという運動です。重力屈性では、茎が重力と反対方向(上)に向かって伸びますが、動物のような感覚器も運動器も持たない植物が、このような複雑な運動をするのは不思議ですよね。
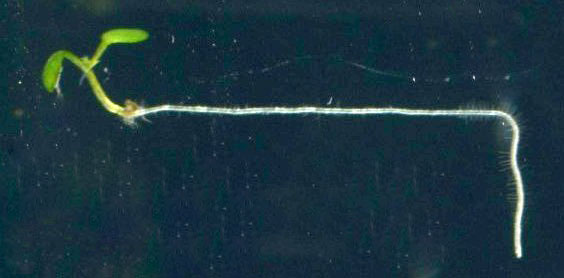
高校生物の教科書に載った写真
重力屈性の仕組みを分子の言葉で理解するために、田坂研究室ではシロイヌナズナを用いて分子遺伝学的な研究が進められていました。重力屈性がおかしくなった変異体を解析して、どの遺伝子が重力屈性に必要かを見つけるのです。私は、シロイヌナズナ花茎の重力屈性異常変異体sgr (shoot gravitropism)シリーズの原因遺伝子の探索を担当しました。まずは表現型を詳しく知るために、シロイヌナズナ花茎の重力屈性をなるべく簡単に観察できるシステムをつくることにしましたが、それには暗黒下で一定時間毎に写真を自動で撮る必要があります。植物が感知できない近赤外光を照明光として用い、試行錯誤しながらCCDカメラでタイムラプス撮影をするシステムを組みました。思った以上に美しい写真が撮れたので、連続写真を重ね合わせて重力屈性の様子がよくわかる写真を作って楽しんでいました。その写真は今、高校生物の教科書(啓林館)に掲載されています。

重力屈性の鍵を握る液胞
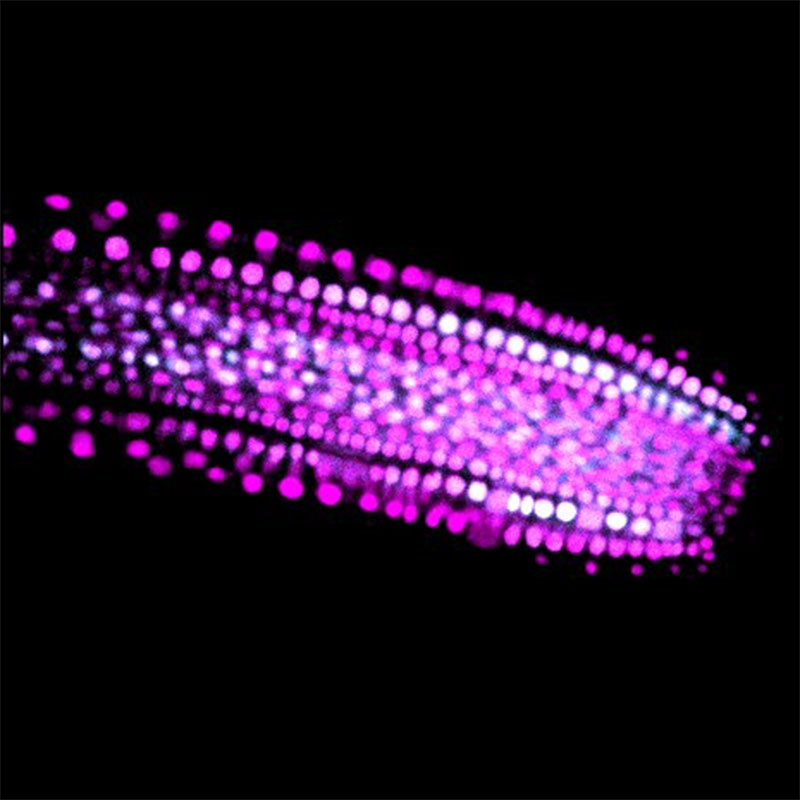
詳しく調べてみると、私が担当したSGR4遺伝子は、細胞内の小胞輸送に関与するSNAREと呼ばれるタンパク質で、液胞へのタンパク質輸送に関与することが分かりました。重力屈性と液胞との関係はそれまでほとんど知られていませんでしたが、植物研究に身を投じて4年目に、花茎では液胞機能が重力屈性に重要であることを明らかにすることができました。ラボメンバーによって重力感受を担うのは茎の少し内側にある内皮細胞であることが示されていたので、内皮細胞の液胞とSGR4との関係を調べ、重力感受細胞の液胞機能が重力屈性の鍵を握ることが見えてきました。
液胞はどの細胞にもありますが、花茎の内皮細胞では液胞が体積のほとんどを占めるほどに発達し、デンプンを蓄積する細胞内小器官であるアミロプラストをぐるりと囲んでいました。アミロプラストの中にはデンプン顆粒が大量に入っていて重いので、重力方向にアミロプラストが沈むことが知られています。重力方向の変化に伴ってアミロプラストが新たな重力方向へ移動する際に、その周りを取り囲む液胞膜がどのようになるのか、興味が湧きました。
世界でひとつだけの顕微鏡
生物の研究で細胞を観察するには顕微鏡を使います。でも普通の顕微鏡では、ステージ上に試料を横たえて観察することになり、重力の方向を変えることはできません。重力応答の研究を地球上で行う難しさはそこにあります。そこで、光学機器メーカーのオリンパス(現EVIDENT)の協力を得て、顕微鏡自体を横に倒してステージを垂直に立て、回転ステージを用いて試料の方向を自由に変えることができる世界でひとつだけの顕微鏡システムをつくりました。

この顕微鏡で内皮細胞を観察してみると、液胞膜はいくつもの陥入構造を持っており、それが形成されたり、位置を変えたり、消失したりと、とてもダイナミックに形を変えることがわかりました。また、植物を逆さまにすると、アミロプラストが重力方向に動くのとともに、周囲を囲むように陥入している液胞膜の形状が自在に変化していく様子が観察されました。ところがsgr4の変異体では、アミロプラストは液胞膜に取り囲まれることなく細胞の縁に追いやられており、さらに驚いたことに、アミロプラストが重力方向に移動することができなくなっていたのです。その後、他の重力屈性変異体でも同様に、液胞機能の異常によってアミロプラストの沈降異常が引き起こされることを突き止めました。細胞が重力を感知するには、液胞機能が正常に保たれ、アミロプラストが自在に動いて重力方向へ沈むことが重要なのです。
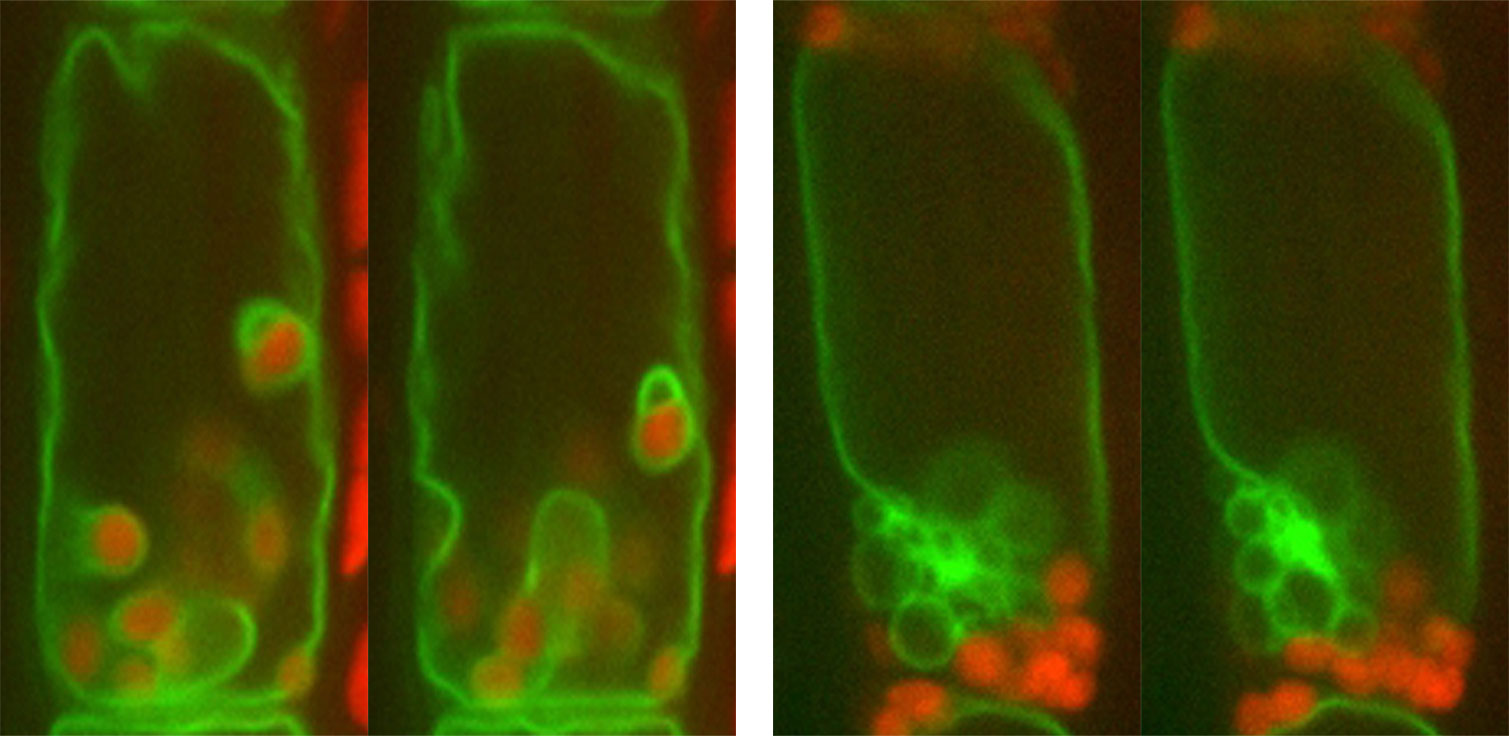
重力感知機構の実体に迫る
アミロプラストが沈む方向をどのように細胞が感知するのか?この重力情報伝達の分子メカニズムが次に知りたくなります。先述したように、花茎では内皮細胞が重力を感知します。そこで、内皮細胞が正しく分化せず重力屈性がおかしくなったシロイヌナズナ変異体と野生株とを比べて、内皮細胞だけで発現し、重力情報伝達に関与する可能性が高いと期待できる遺伝子群を絞り込むことにしました。絞り込まれた上位3つの遺伝子は互いに少しずつ似ていて同じ遺伝子ファミリーと考えられましたが、当初はその機能が全く分かりませんでした。しかしあるとき、このファミリーに似た遺伝子が、イネの重力屈性異常変異体lazy1の原因遺伝子として報告されていることに気がつきました。似ていると言ってもほんのわずかで、専用の検索ソフトでも見つからないくらいでしたが。
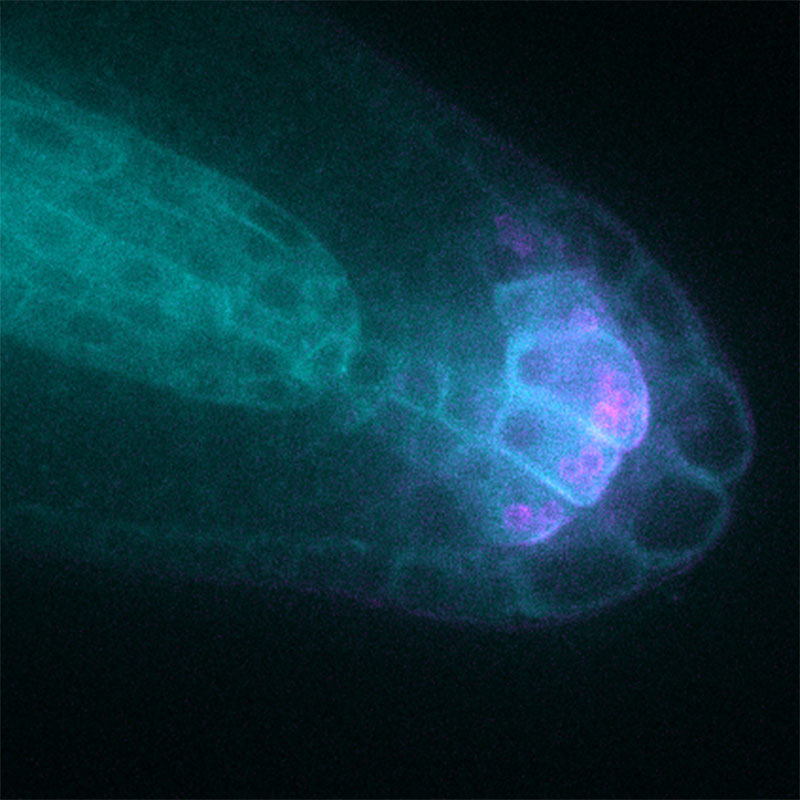
そこで、絞り込んでいたシロイヌナズナの遺伝子ファミリーにLAZY1-LIKE (LZY)と名前をつけ、詳しく調べることにしました。顕微鏡で丁寧に観察してみると、LZYタンパク質は普段はアミロプラストに存在するのですが、面白いことにアミロプラストが重力方向に沈んで細胞膜に近付くと、LZYタンパク質が速やかに細胞膜上に移動することを発見しました。つまり、LZYタンパク質は、アミロプラストの位置情報、すなわち重力方向を細胞膜に伝達する情報分子として働く重力感知機構の実体だったのです。こうして、植物の重力屈性のしくみに決定的な分子モデルを提案することができました。
ワクワクする基礎研究が社会につながる
私にとっての研究の醍醐味は、「帆船の見張り台から誰よりも早く大陸を見つける」ような経験をすることです。足場は悪いし、風に煽られるかも知れませんが、「何か新しいものを見つける」とゾクゾク、ワクワクするじゃないですか。私が最初にその経験をしたのは、大学院生の時です。オートラジオグラフを暗室の安全光に照らして、はっきりとバンドが見えた時、思わずガッツポーズをしました。LZYタンパク質が重力に応答する瞬間を、自分が構築した垂直ステージ共焦点顕微鏡で捉えることができた時も、大陸を発見した気分でした。さすがにもうガッツポーズはしませんでしたが、ラボメンバーと大いに盛り上がりました。
重力屈性に異常が生じると、植物が倒れたり、根や枝が横に広がったりします。このような性質は収穫や密植がしにくくなり収量が落ちるので、農業においてはあまり歓迎されません。つまり、重力屈性が関与する枝や根の成長角度は、生産量に直結する重要な形質として注目されてきたのです。これまでに、イネをはじめとしてトウモロコシ、マメ科植物、コムギ、ソバ、セイヨウアブラナ、バラ、リンゴなど、多くの作物で、LZYが枝や根の角度(重力屈性)の決定に重要な働きを持つ遺伝子として単離されています。私たちの分子レベルでの基礎研究が、この様な収量に関与する作物の形質に深い理解をもたらし、将来的に作物の生産性を上昇させることに繋がれば嬉しいです。

構成協力/撮影 松林 嘉克
森田 美代 プロフィール:
1990年 愛媛大学理学部卒業。京都大学大学院理学研究科にて博士(理学)の学位を取得。(株)HSP研究所研究員、奈良先端科学技術大学院大学助手、同准教授、名古屋大学農学部教授を経て、2018年より現職。