本来独立栄養生物であるはずの植物が、寄生植物として従属栄養生物になる生存戦略を選んだ例が多々ある。それは何故なのか、そしてどうやってそれをなしとげたのか?また、どうやって寄生植物が別の植物を認識し、寄生を開始することができるのか?この不思議に魅了され、寄生植物の分子生物学的研究を始めた研究者がいる。そのうちの1人、理研環境資源科学研究センター・植物免疫研究グループの白須賢グループディレクターは、独自の研究システムを開発し、寄生に重要な遺伝子群を単離してきた。それまで植物免疫分野を中心に研究を推進してきた白須グループディレクターに、どうして寄生植物の研究を始めたのか、そして何が分かってきて、さらに何を明らかにしていきたいのかを聞いた。
小学校の夢は科学者
小学校4年ぐらいの頃だと思います。実は記録があって、小学校4年生の文集に将来なりたいものとして科学者(生物学者)と書いているのです。“動物や虫などがすきで大すきだから(ファーブルのように)“とあります。よっぽど好きだったのですね。虫網を持って走り回っていました。お花を見るのも大好きでしたよ。山で見つけた赤い実を小学校の校長先生の所に持っていってこれは何の実ですかと聞いたこともありました。その校長先生はびっくりしたでしょうが、ちゃんと調べて教えてくれました。とても嬉しかったのを覚えています。

大学では分子生物学をやりたくて、農学部に進みました。植物の分子生物学をやろうと、植物ホルモンの大先生のところにお伺いしてお話を聞くと、うちでは生化学が主でホルモンの単離はやっているが、分子生物学はやっていない。ここで分子生物学をやりたいなら微生物をやった方がよいだろうと言われました。それならと分子生物学をやっている微生物の研究室に入りましたが、与えられた研究テーマは、膜タンパク質の単離精製でした。それはそれで楽しく、いろいろな技術を学ばせてもらったのですが、結局、大学院からはアメリカの植物研究が強い大学に留学しました。それからはずっと植物と病原体の相互作用についての研究をしています。
留学先で寄生植物と出会う
当時、私が所属していた英国の研究所は植物の免疫をテーマに多くの研究者が集まるハブとなっていました。そこには、アフリカ出身の研究者も何人かいて、いい友達になったひとりが、何故、誰もアフリカで大きな問題になっている寄生植物ストライガの研究をしないのだろうと話していて、この植物を初めて知りました。しばらくたって、別のアフリカ出身の研究者である友人から、ストライガのグラント(研究助成金申請書)を書いたから見てくれないかと頼まれました。アフリカからストライガを集めてその多様性を研究するというもので、とても魅力的なものだったことを覚えています。結局、彼女はそのグラントの獲得に成功しストライガの研究を始めるのですが、逆にその業績を買われ、とある財団にヘッドハンティングされてしまい研究は頓挫してしまいました。

その頃、私は縁があって理研に移ったのですが、理研のような研究環境でしかできないような研究を始めたいと思い、ストライガの研究ができないかと考えたのです。研究内容は、寄生の分子メカニズムです。これは誰もやっていませんでした。ストライガの研究そのものが難しかったのですね。まず初めにしたことは、この研究をやってみたいと思う人を探してくることでした。システムのセットアップもほぼゼロからで、おそらくなにか面白いことが分かるのに10年ぐらいかかるだろうけど、それをやってくれる人がいるのだろうかと当初は不安でした。しかし、英国で同じ研究所で研究していた吉田聡子さんが、既にドイツで助教をしていたのですが、帰国して参加してくれることになりました。
寄生の鍵分子ストリゴラクトン
ストライガは植物が根から分泌する化合物であるストリゴール(ストリゴラクトンと呼ばれる化合物の一種として初めて同定されたもので、名前はストライガからとっている)を認識しないと発芽しません。ストリゴールは土壌中ですぐ分解されてしまうので、寄生植物にとっては、宿主の根がすぐ近くにあるという証拠なのですね。とにかく、最初にストリゴールを手に入れる必要がありました。しかし、天然のストリゴールは微量にしか存在せず、しかも不安定、さらに有機合成は立体異性体を生ずるために非常に難しいので、製品として売っていなかったのです。幸運にも、引退されていた大学時代の有機化学の先生である故・森謙治先生がまだ合成した化合物をお持ちで、一升瓶をもってご自宅にお伺いして一緒にお酒を飲むと、なんとそのすべてを譲っていただけました。これで実験ができるようになったのです。
陸上植物の80%は菌根菌と呼ばれるカビの一種と根で共生しています。菌根菌は、土中にも菌糸を張り巡らし、植物の根が届かない場所のリンなどを植物に届けることができるのですが、その菌根菌を根に呼び込むために、多くの植物が根からストリゴラクトンを分泌すると考えられています。また、内生のストリゴラクトンは、植物の脇芽の発生を阻害する機能があることが分かりました。これは、森先生からいただいたストリゴールがあったからできた実験で明らかになったものです。この後、ストライガにおけるストリゴールの受容体も明らかになりましたが、この発見には、吉田さんが中心におこなったストライガのトランスクリプトームやゲノムの解析が役に立ちました。驚くべきことに、ストライガはストリゴラクトンの受容体をコードする遺伝子を20個近く持っていたのです。普通の植物では1つか2つですので、ストライガがストリゴラクトンの受容に関して大きな進化を成し遂げていたのです。
寄生研究のモデル植物を求めて
ストライガのストリゴラクトン受容体遺伝子の解析は、モデル植物のシロイヌナズナを使って行なわれました。一方、ストライガは絶対寄生植物なので、育てるのに宿主が必要であるばかりか、形質転換の手法や、遺伝子の遺伝学的同定手法も確立できていません。なので、急がば回れと、寄生植物のモデルを構築することにしました。条件は、ストライガと同じハマウツボ科の日本在来種で、遺伝学がしやすいように自家受粉で2倍体、かつ宿主がなくても生きていける条件的寄生植物です。こんな植物がないかなと探していた時に、縁あって軽井沢町植物園を訪れました。名誉園長の故・佐藤邦雄先生(当時94才!)が、それならコシオガマを見せてあげようと、わざわざご自分で案内してくれました。数ヶ月したら種がつくから送ってあげるとまでおっしゃってくれたのです。

最初は、水が合わなくて枯れてしまったりして、育てるのが難しかったのですが、日本の各地から種をとりよせたり、少しずつ種を増やして、なんとか研究ができるようになりました。ブラジルからの大学院生イシダ・ジュリアーニさんが形質転換の系を立ち上げてくれて、機能解析もできるようになりました。結局、変異体を使った遺伝学的実験の論文が出るまでに10年ぐらいかかりましたね。そしてトランスクリプトームそしてゲノムもシークエンスして、寄生植物がもつ遺伝子群の全容が明らかになりました。
コシオガマが開いた突破口
これまでの研究でハマウツボ科の植物は、宿主植物の細胞壁由来のキノン化合物を認識すると寄生に必要な器官である吸器を根の先端に作ることが分かっていました。このキノン化合物は土壌中で不安定であるが故に、寄生植物にとっては、そこに生きた宿主植物がいる証拠なのでしょうね。なので、コシオガマにこのキノン化合物をかけて、すぐに発現上昇する遺伝子群を調べました。もちろん多くの遺伝子の発現が上昇するのですが、最初に私たちが注目したのは、植物ホルモンであるオーキシンの生合成にかかせない酵素をコードする遺伝子でした。この遺伝子は、これから吸器ができる場所の表皮細胞で発現が上昇します。オーキシンの蓄積を示すマーカーもまず吸器の先端でみられました。オーキシンは細胞分裂を誘導するので、この部位で根が膨らんでくるのも納得できます。面白いのは表皮に既に分化していた細胞が新しい機能を持った細胞に再分化していくことです。そしてこの細胞群がまるで目があるかのように宿主の細胞間隙に入り込み、その維管束に向かっていきます。いったん宿主の道管に達すると吸器の内部に自身の道管を形成してつなぎます。
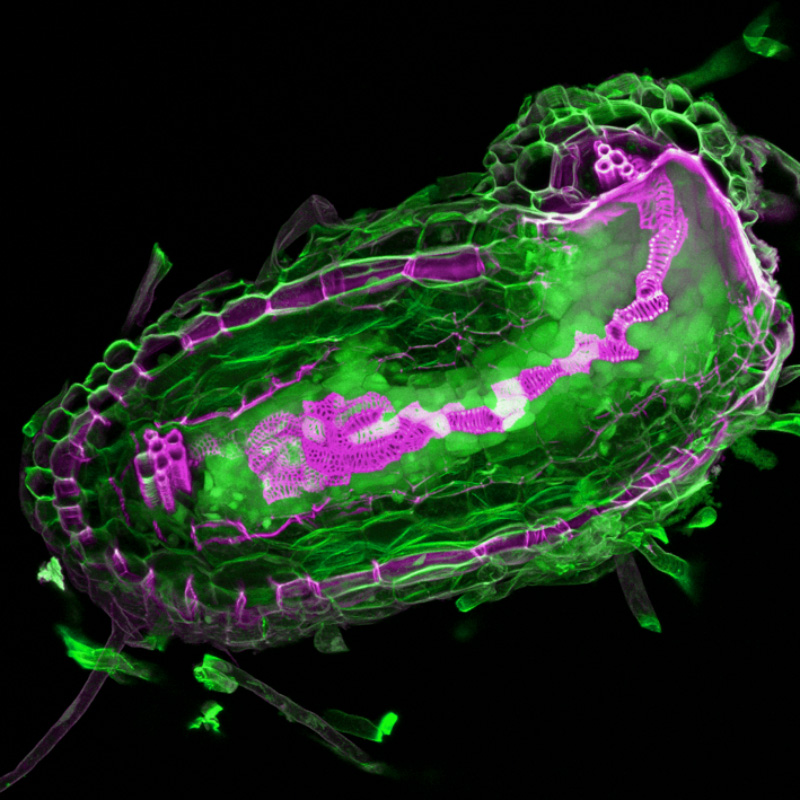
寄生植物が宿主植物を見つけるしくみ
最初に考えたのは、どうやってハマウツボ科の寄生植物はキノン化合物を認識できるようになったのだろうか、ということでした。そしてそれをどうやったら解明できるのかと考えていました。しかしある時、もしかしたらこの質問そのものが間違っているのではないか、もしかしたらすべての植物はキノン化合物を認識できて、ハマウツボ科の植物だけがそのシグナルを寄生に使うようになったのではないかと考えたのです。調べてみると寄生植物ではないシロイヌナズナもキノン化合物を感知していることが分かりました。そこでシロイヌナズナでキノン化合物を認識できない変異体を多数単離して、ゲノム上でどの遺伝子が壊れているかを探したところ、すべての変異体で、ある受容体様タンパク質をコードする遺伝子が壊れていることが分かりました。コシオガマもこの遺伝子の相同遺伝子をもっていて、そのいくつかが根の表皮細胞で特異的に発現していました。また、その遺伝子をシロイヌナズナの変異体に導入するとキノン化合物を認識できるようになりました。この遺伝子はコケにも存在することから、キノン化合物を認識することは陸上植物に普遍的な現象だと考えています。
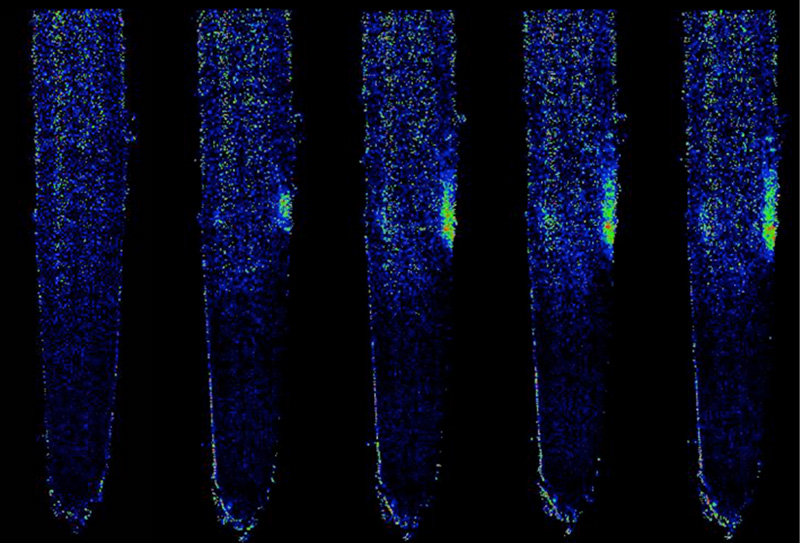
他者認識の普遍的メカニズム
キノン化合物を認識できないシロイヌナズナの変異体でキノン化合物をかけたときの遺伝子発現解析をしたところ、傷や病原体に感染したときに発現する遺伝子群の発現が落ちていました。私の研究室のメインテーマは植物の免疫ですから、すぐにピンときました。研究室にある病原細菌をその変異体にかけてみると、それに対する免疫が落ちていることが分かりました。その病原菌は気孔から侵入してくるのですが、この変異体では気孔の開閉がおかしくなっていました。つまり、シロイヌナズナはキノン化合物を認識すると気孔を閉鎖し、病原菌に抵抗していたのです。この場合、キノン化合物が病原菌によって壊された細胞壁からできるのか、病原体由来のキノン化合物なのかは、まだ分かっていません。いずれにせよ、植物が他者を認識するメカニズムのひとつだったということですね。
面白いことはその辺に転がっている
これまで分かってきたことは、まだほんの少しだと思っています。例えば、ストライガやコシオガマがキノン化合物を認識して吸器を作る前に、宿主に向かって根を伸ばしていくのですが、その機構は明らかになっていません。この事象は寄生の最初のステップであり、進化において非常に重要な鍵を握っていると思って研究を進めています。また、キノン化合物を認識したあと、宿主に一番近い表皮のオーキシン濃度が上がるわけですが、そこに至るシグナル伝達系も分かっていません。感染した後、寄生植物と宿主植物の間でどのようなコミュニケーション、あるいは核酸情報の移動があるのか、とても興味のあるところです。キノン受容機構の研究のように、寄生植物というちょっと変わった研究材料を用いることで、植物の普遍的な生物学的メカニズムが見えてくるかもしれません。面白いことはその辺に転がっているということですかね。画家のアンリ・マティスもこう言っています。”見たいと思う人には,どこにでも花はあるのです” 。これからもいろんな花を見ていきたいですね。
構成協力/撮影 松林 嘉克
白須 賢 プロフィール:
1988年 東京大学農学部卒業。カリフォルニア大学デービス校にてPhDを取得。ソーク研究所ポスドク、セインズベリー研究所グループリーダー、理化学研究所植物科学研究センター(現・環境資源科学研究センター)グループディレクターを経て、2020年より現職。