窒素は生物が生きていくために最も重要な栄養素と言っても過言ではない。窒素栄養を自分でつくることができない植物は、窒素栄養を得るために様々な戦略を発達させている。その最たる例は、窒素固定細菌・根粒菌との共生ではないだろうか。根粒共生と呼ばれる相利共生現象であるが、パートナーがお互いを認識し、共生器官の根粒が形づくられてそれが機能をもつまでには、二者間での巧妙な相互作用のしくみが存在している。根粒共生は環境の変化に大きく影響を受けることが知られているが、共生と環境、一見無関係にみえる2つの事象が壽崎拓哉准教授のグループの研究によってつながり、150年来の謎もそのしくみが解き明かされつつある。微生物との共生をヒントにして植物の環境適応のしくみの理解に迫る壽崎准教授に、研究の魅力や今後の研究展開について聞いた。
はじまりはイネから
植物を使って人の役に立ちそうなことを研究してみたいと漠然と考えていたこともあり、農学部に進み、平野博之先生の研究室で、イネの分裂組織(メリステム)の維持機構に関する研究に取り組みました。メリステムは茎の先端にある未分化な細胞から構成される組織で、葉や花などの器官をつくるために重要な役割をもっています。学部4年生の時に最初に与えられたテーマは、イネのfloral organ number 1 (fon1)変異体とその原因遺伝子の解析でした。

fon1変異体は雌しべや雄しべなどの花の器官数が増加するという表現型を示し、それは花器官をつくる花メリステムのサイズが増大していることが理由でした。一般的に遺伝子の機能はその遺伝子が機能を失った変異体の表現型から推測することができます。fon1変異体の表現型からFON1 遺伝子は花メリステムのサイズが大きくならないようにする機能をもつということが分かりました。
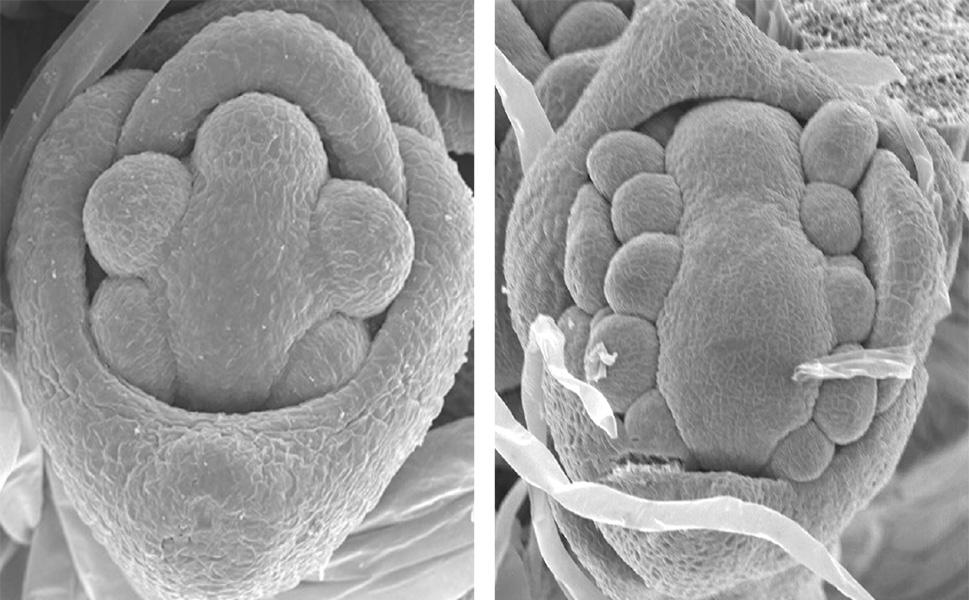
FON1遺伝子がコードしているタンパク質は、受容体型キナーゼの一種でした。受容体型キナーゼは細胞の外で低分子のタンパク質などをリガンドとして受容し、細胞内へと情報を伝える働きをもっています。当時研究の進んでいたシロイヌナズナでは、茎頂メリステムの維持に重要な働きをもつCLV1というタンパク質が知られていました。FON1とCLV1はタンパク質の構造がとてもよく似ていることが分かったことから、植物種が異なっても茎頂や花のメリステムを制御する情報伝達のしくみは基本的には保存されていることを示すことができました。その後の研究により、FON1のリガンドの候補を発見し、また、イネに特有の情報伝達のしくみがあることも分かり、メリステムを制御するしくみの多様性の理解に繋がりました。
FON1の研究を通して、植物の形づくりの研究の面白さを知り、応用的な研究よりも基礎的なしくみを追求する研究を行いたいと考えるようになりました。大学院の途中で平野先生が理学部に異動することになりました。その時には何学部かは関係なく、今の研究を続けることを優先させたいと考えるようになっていましたので、平野先生と一緒に理学部に移り、メリステムの研究を続けました。
私たちがFON1の研究を進めていた頃、近所の川口正代司先生(現・基礎生物学研究所教授)の研究室ではマメ科植物のミヤコグサを使って植物と根粒菌の共生の研究が行われていました。川口先生によってマメ科植物ではHAR1というFON1とよく似た受容体型キナーゼが、茎頂メリステムの制御ではなく根粒菌との共生器官である根粒の数を制御するという論文が発表されたのもちょうどその頃です。FON1とHAR1がタンパク質の構造がよく似ているということがきっかけで植物と根粒菌の共生の世界を知ることになります。博士号を取得後、海外にポスドクとして留学しメリステムの研究を続けていたのですが、今後どんな研究をしていこうかずっと考えていました。そんなある時に、川口先生から「植物と根粒菌の共生の研究に興味はないか?」とアカデミックポストのお誘いの連絡をいただきました。

メリステムから共生の研究へ
今後も植物の形づくりの研究は続けたいと考えていましたが、これまで進めてきたものとは毛色の異なる現象を対象にしたいと考えていたこともあって、川口先生の研究室にお世話になることを決めました。川口研究室では、根粒形成に関することであれば研究テーマは割と自由に決めることができましたので、どんな研究をするか考えるところから始まりました。研究の醍醐味はまだ世界で誰も知らないことを明らかにすることにあると考えています。したがって、研究を始める前の予備知識として何が分かっていて、分かっていないのかを把握しておく必要があります。また、分かっているようでいて実は分かっていないことも多くあります。これらのことを判断するための最も有益な情報源は論文です。論文をたくさん読み、植物と根粒菌の共生を題材にして何の研究をするかよく考えました。
植物と根粒菌の巧妙なやりとり
マメ科植物は根粒を根につくり、その中で根粒菌を住まわせることで根粒菌が大気中から固定した窒素を栄養として受け取り、代わりに光合成産物である有機物(炭素栄養)を根粒菌へと提供します。植物の器官形成が微生物との相互作用によって始まって、その器官が窒素固定を行うための特別な機能を発達させていくというところが根粒共生の特徴です。ここでは、ホルモン、代謝産物、タンパク質などの情報分子のやりとりのことを相互作用ということにします。根粒形成を開始するために、植物はフラボノイド、根粒菌はリポキチンオリゴ糖の一種というように特定のシグナル分子を土中に放出し相手に認識してもらいます。シグナル分子の認識後、植物は根の表皮に存在する根毛をくるっと曲げて根粒菌を捕まえつつ、根の皮層細胞が脱分化し根粒をつくるための細胞分裂を始めます。また、植物は感染糸とよばれる根粒菌の通り道を形成し、根粒菌に感染糸を介して根粒細胞の中に入ってきてもらいます。
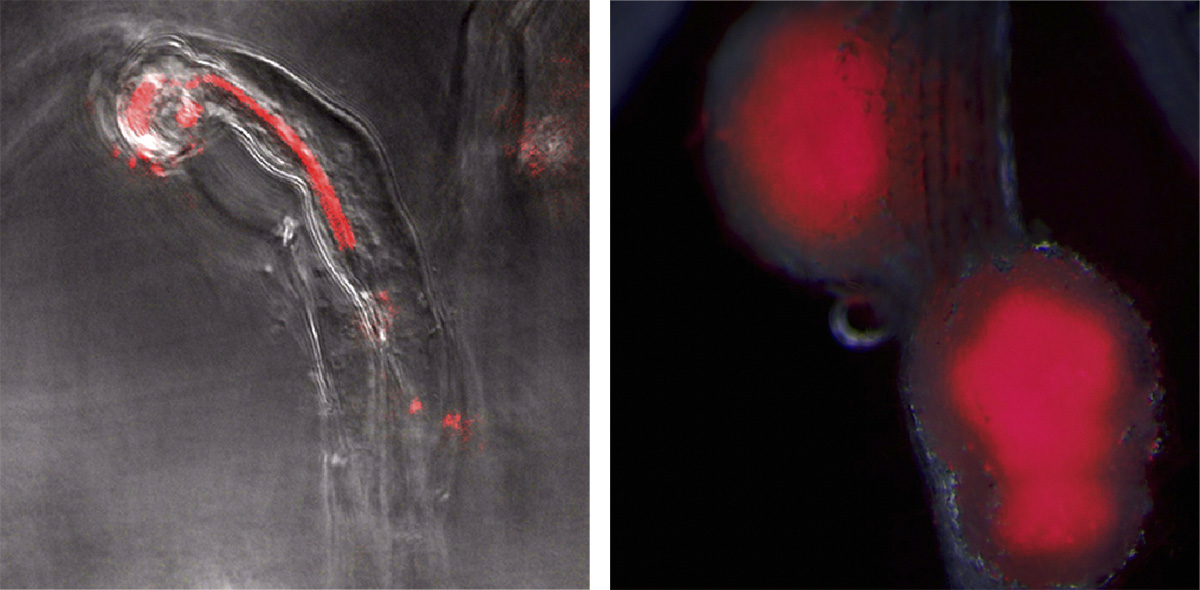
根粒内部に細胞内共生した根粒菌はニトロゲナーゼと呼ばれる酵素を用いて窒素固定を行います。マメ科植物と共生するタイプの根粒菌は、土中に単独で存在するいわゆるfree livingの状態では窒素固定を行うことができません。それは、ニトロゲナーゼが働くために必要な補助因子を、根粒菌自身ではつくることができないからです。この補助因子は根粒菌が根粒の中で細胞内共生しているときに植物から提供されます。根粒菌は窒素固定をする能力をもっているのに、植物の助けがなければ窒素固定がすることができないなんてまさに相利共生という感じがしませんか。
共生のコストとメリット
先ほど話したように、根粒器官の形成、根粒菌の感染、窒素固定などいくつかのイベントが連続して起こることによって根粒共生が成立します。私の研究室ではこれら個々のイベントの制御に関わる植物の遺伝子の研究を行っていますが、現在、力を入れて取り組んでいる研究は根粒共生と環境の関わりです。
根粒共生は窒素栄養の不足した土壌で植物が生育する上では非常に有用です。しかし、根粒共生は植物にとって必ずしも良いことばかりではないのです。植物は根粒共生によって窒素栄養を得ることができる一方で、根粒の器官形成や根粒菌の窒素固定反応には光合成で作られた炭素栄養がエネルギー源として使われます。根粒共生に炭素栄養が使われすぎると植物は自身の成長につかう炭素栄養が少なくなってしまい、うまく成長することができなくなってしまいます。植物は根粒由来の窒素の獲得というメリットと炭素栄養の消費というコストのバランスをうまく保ちながら根粒共生を行なっているのです。

土壌中に窒素栄養が豊富に存在する土壌では、植物は根粒共生を行わなくても窒素栄養を直接土から得ることができます。そこで植物は、このような環境では根粒に依存した窒素栄養獲得戦略から土壌中の窒素栄養を直接得る戦略に切り替えることで、根粒共生に伴う不要なエネルギーの消費を防いでいます。この「植物は窒素栄養の豊富な環境下では根粒共生をやめる」という現象は150年ほど前から知られている現象なのですが、そのしくみはほとんど分かっていませんでした。
この研究では、まずこのしくみに関わる遺伝子を探すところから始めました。このしくみに関わる遺伝子がこわれた突然変異体では高濃度の窒素栄養が存在している土壌でも根粒を形成するのではないかと予想し、そのような表現型を示す突然変異体を見つけようと考えました。その仮説は正しかったようで、変異体スクリーニングによってnitrate unresponsive symbiosis (nrsym) と名前をつけたこれまで報告例のない突然変異体をいくつか単離し、さらに原因遺伝子を特定することができました。

そのなかのNRSYM1遺伝子は転写因子と呼ばれるDNA配列に特異的に結合し、標的となる遺伝子の発現を調節する機能を持つタンパク質をコードしていることが分かりました。NRSYM1転写因子の働きに着目した研究によって、窒素栄養が豊富に存在する土壌では根だけでなく地上部も介して根粒形成がコントロールされることや根粒形成に関わる遺伝子の発現が調節されるしくみが分かってきました。150年来の謎が私たちの研究によって少しずつ解き明かされていることを実感しています。
解けた疑問と残された謎
今後も、動くことのできない植物が芽生えたその地で過酷で様々な環境ストレスに晒されながらもたくましく生きて子孫を残している、その環境適応のしくみを根粒共生の研究を通して分子レベルで深く理解していきたいと考えています。私たちが研究している「植物は窒素栄養の豊富な環境下では根粒共生をやめる」という現象は栄養に対する植物の応答がわかりやすく観察されるので、植物の栄養環境適応を研究する上で良い材料になると考えています。
根粒共生は植物が窒素栄養を上手に手に入れるために植物が進化させてきた環境適応戦略の1つです。マメ科植物は根粒と窒素栄養の獲得に特別化した器官をつくるようになったおかげで、根粒をつくる能力をもっていない植物では生きられないような貧栄養な土地でも生育することができます。それでは、なぜ、多種多様な植物のなかで主にマメ科植物だけが根粒形成を行うことができるのでしょうか。これは根粒共生の研究の中で最も大きな問いのひとつだと思います。現在は遺伝子の進化の観点から少しずつですがその理由も分かってきましたが、未解明な点が多く残されています。植物の進化の過程で根粒形成がどうやって生まれたのか、その進化基盤を明らかにできれば、その知見を活用してマメ科以外の植物にも根粒をつけさせることができる可能性があります。もしそれが実現できれば、植物がこれまで育てられなかったような貧栄養な土地での植物の栽培にもつながることから食料問題の解決に大いに貢献できるのではないかと期待しています。
パズルのピースを探して
学部4年生の時に研究室に配属されてから、研究が面白くて続けていたらいつのまにか研究を仕事にしていました。初めはなんとなく植物の研究を選んだのかもしれませんが、今では環境に適応するために進化した植物独自の緻密なシステムに魅了されています。
研究には終わりはなく、進めば進むほど新たな疑問がわいてきて、その疑問を解くための次の研究が始まります。また、研究はうまくいくことよりもうまくいかないことの方が多く、思うように進まず悩むことも多いです。予想と実験結果が異なる時は、立てた仮説が間違っているのか、実験のやり方がよくなかったのか等、必ず理由があります。その理由に気づき真相に辿り着けるようにもっと考えたり、条件を検討してさらに実験を繰り返します。ああでもないこうでもないとあれこれ考えそれを実行する過程も案外楽しんでいるのかもしれません。また、研究を続けていると、ごく稀に頭の中のパズルの最後のピースがピタッとはまるような、すごくよいと思えるデータを得ることがあります。そんな時はそれまでの苦労も一気に吹き飛びます。そんな瞬間にまた出会いたくて、これからも研究を続けていくのだと思います。
構成協力/撮影 松林 嘉克
壽崎 拓哉 プロフィール:
2003年 東京大学農学部卒業。東京大学大学院理学系研究科にて博士(理学)の学位を取得。日本学術振興会特別研究員、基礎生物学研究所助教を経て、2015年より現職。