動き回ることのできない植物は、周囲の環境を鋭敏に感じ取り、その変化に動物以上に頑強かつしなやかに適応する能力を進化させてきた。なかでも植物の成長に最も影響を与える光環境の変動に対しては、フィトクロムと呼ばれる光受容体を中心とした精緻な光情報認識機構を進化させ、極めて巧妙に適応するしくみを持っている。フィトクロムは1959年に発見されて以来、世界各国で精力的に研究が進められ、「フィトクロムの研究はもうやり尽くされてしまったのではないか?」と言われて久しい。そのような研究分野において、松下智直教授のグループはこれまでに次々とブレークスルーを生み出してきた。どのような背景でそれらの発見がなされてきたのか、また何をモチベーションに研究を行っているのか、飾らぬ実情について尋ねてみた。
植物に囲まれて育った
祖母と母が園芸好きだったので、たくさんの植物に囲まれて育ちました。庭に育っている様々な植物を眺めたり、小学校の頃に両親に買ってもらった顕微鏡で植物を観察したりするうちに、その内面にも興味が湧いてきて、植物の中にミクロの広がりを想像することが大好きになりました。友達には内緒でしたが、盆栽の収集や栽培に熱中していた時期もあります。盆栽には自然から切り取られた一場面のエッセンスが一鉢の中に再現されている気がして、その中に自然風景の広がりを子供ながらに感じていました。そんな少年時代を過ごすうちに、いつの間にか、将来は植物の研究者になりたいと思うようになっていました。
フィトクロムの機能美
植物はフィトクロムという光受容体タンパク質を持っていて、自然光に含まれる赤色光成分と遠赤色光成分の比を感知して、様々な適応反応を示すということを、高校の生物の授業で知りました。光合成では主に青と赤の光が使われるので赤色光成分はとても重要なのですが、他の葉の陰に入ってしまうと青と赤の光は減って遠赤色光が届くので,植物は赤色光成分と遠赤色光成分の比で自身の光環境をモニターしているのです。この時、フィトクロムは赤色光を吸収して活性型へ、また遠赤色光を吸収して不活性型へと可逆的に変化します。何と合理的で洗練されたシステムが植物細胞の中には備わっているのだろうと思いました。

その後大学に入って勉強を進めると、ますますフィトクロムの機能美に惹かれていきました。フィトクロムが試験管の中で示す光化学的な性質から、例えばどのぐらいの赤色光と遠赤色光をどういうタイミングで照射すれば、活性型フィトクロムがどれぐらい生じるかというようなことが計算できるのですが、その計算結果が、植物を実際にその光にさらしたときに観察される応答の量にほぼそのまま反映されるのです。複雑でファジーなしくみを持った生物において、物理化学的法則がそのままきれいに反映される応答系は希なので、フィトクロムを介した植物の光に対する応答系の美しさが特に際立って感じられました。
学説に反する意外な結果
本格的にフィトクロムの研究に携わることになったのは大学院に入ってからだったのですが、期待に反して最初の3年間は、ポジティブデータが1つも出ないという絶望的な状況でした。博士課程1年の秋頃は、自分の生産性の低さに嫌気がさして不登校になるほどでした。大学には行かず、近所の飲み屋を転々とする毎日でしたね。
フィトクロムの実体はタンパク質で、その1分子は、N末端側部分とC末端側部分の2つの部分に大きく分けられます。当時フィトクロムは、N末端側部分で光を受け取り、そしてもう一方のC末端側部分でその情報を別のタンパク質に伝えることで、植物の光応答を引き起こすと信じられていました。フィトクロム研究の歴史は比較的長く、1959年に発見されて以来、研究データの膨大な蓄積があります。それらの多くが上記の学説を一見支持するかのように見えたので、当時この分野でその説を信じない者は一人もいなかったのではないかと思います。もちろん当時の私もそれを100%信じ込んでいました。しかし、その説に基づいて研究計画を立て実験を行っても、期待通りのデータが1つも出ませんでした。
でも博士課程1年の終わり頃に転機が訪れました。フィトクロムのN末端側部分のみを発現する植物が赤色光に対してわずかに応答を示すという、これまでの説に反する意外な結果を得たのです。最初は半信半疑だったのですが、その結果は何度も再現されました。そこからは、やればやるだけ面白いデータが出続けて、論文発表までにそれほど時間を要しませんでした。この成果によって、細胞内でフィトクロムがC末端側部分からではなく、N末端側部分から光情報を発信していることが明らかになりました。
また、フィトクロムがC末端側部分の働きによって二量体化し核移行することの意義についても明らかにすることができました。C末端側部分は、光情報の発信に直接関わらないばかりか、むしろそれに対して阻害的に働いていました。これら一連の発見により、フィトクロム分子内の構造と機能に関する様々な謎が一気に解消されるとともに、これまでの学説が完全に覆され、フィトクロムの細胞内での機能について一から考え直す必要が生じたのです。こうなると、俄然として研究が面白くなってきました。

留学先でワインを学ぶ
小さい頃から、植物科学の研究を海外で行うことが夢だったので、留学先選びには力を入れました。留学先の研究内容ももちろん重要ですが、それと同じぐらいその地方の文化、特に飲食の文化に興味があったので、ワインの産地の中から留学先を選ぶことにしました。そうして決めたのが、アメリカ西海岸にあるUCLA(カリフォルニア大学ロサンゼルス校)でした。
期待通り、ロサンゼルスでの西海岸生活は大変楽しく、充実したものでした。特にワインと肉料理について体系立てて勉強できたことは、一生の財産となりました。一方、UCLAでの研究テーマは、植物の青色光受容体であるクリプトクロムの細胞内での機能を明らかにしようとするものだったのですが、これがまるでうまく進みませんでした。結局、2年間の留学で論文を1本も出すことができませんでしたが、その間に考え貯めた実験アイディアの蓄積は確実に今に活かされていますし、何よりも、家族とともにロサンゼルスで過ごした2年間の経験は、その後の人生を何倍にも豊かにするものだったので、留学は大成功だったと思っています。

誰もやっていないことを研究したい
帰国後、九州大学で自分の独立した研究室を持つことができました。好きな研究をできるせっかくのチャンスなので、それならば誰もやっていないことを研究したいと思い、フィトクロムに関する新たなテーマをゼロから探すことにしました。そこで改めて大規模な変異体スクリーニングを行って、フィトクロムの働きに必要な新しい遺伝子を探すことにしたのです。変異体スクリーニングの魅力は何と言っても、ひとつひとつの観察をしっかり行っておきさえすれば、確実にその現象に関わる遺伝子が同定でき、しかも全く予想もしないような遺伝子に辿り着ける可能性があるという点です。
この試みは成功し、選択的スプライシングの制御に関わるタンパク質がフィトクロムの働きに必要であることを発見することができました。選択的スプライシングとは、DNAから転写されたmRNA前駆体の中から、異なる組合せのイントロン配列がスプライシングにより除去される現象のことで、1つの遺伝子から配列の異なる複数のタンパク質を生み出すしくみのひとつです。それまでフィトクロムは、様々な遺伝子の転写量を制御することで光応答を引き起こすと考えられていたのですが、その概念を一新させるような成果になりました。

スプライシングを制御するフィトクロム
こうなると、フィトクロムの働きに伴って選択的スプライシングの変化を示す遺伝子がどれだけの数存在するのかを網羅的に調べたくなりますよね。でもそれを行うためには、次世代シーケンサーによるmRNA配列の大規模情報解析技術が必要でした。非常に幸運なことに、ちょうどその時期に、情報解析のプロである花田耕介さんがご近所の九州工業大学に異動して来られ、この出会いが私の研究スタイルを大きく変えることになりました。そして共同研究の結果、フィトクロムが転写量制御に加えて、ほぼ同じ規模で選択的スプライシング制御も行うことで、光応答を引き起こしていることを示すことができました。つまりフィトクロムは、遺伝子発現の中の転写とスプライシングという2つの過程を同時に制御することで、mRNAをゲノム規模で量的にも質的にも変化させ、光に対する植物の適応反応を誘導していることが明らかとなったのです。
研究は予想外の展開へ
フィトクロムによる選択的スプライシング制御を受ける1,500以上の遺伝子の中から1遺伝子だけ、グリセリン酸キナーゼ(GLYK)遺伝子を代表例として選び、その制御の生理学的意義とメカニズムについて集中的に解析していきました。当時、博士研究員1名、大学院生1名、そして技術補佐員2名しかいない、とても小さな研究室だったので、対象を極限まで絞って集中的に研究を進めないと世界に太刀打ちできないという事情がありました。このプロジェクトの担当だった博士研究員の牛島智一さんに、「直感でいいから1遺伝子だけ選んで下さい、今後はそれに集中しましょう」と伝え、そして牛島さんが選んだのがGLYK遺伝子でした。
しかしGLYK遺伝子に絞った研究を始めて2年ほどが経った頃に、フィトクロムの働きによって赤色光依存的にGLYK遺伝子の転写開始点(遺伝子の転写が始まる場所)が変化し、それによって直後のイントロンの始まる場所がずれているということが分かりました。つまり、フィトクロムによるGLYK遺伝子の選択的スプライシング制御は間接的なものだったのです。これは相当ショックでしたね。2年間で作り上げてきた形質転換植物やその他のリソースが、ほぼ全て無駄になってしまったわけですから。1,500以上もある遺伝子の中から、よりによってなぜGLYKなんかを選んでしまったのかと、自分たちの引きの悪さを呪いました。
1週間ほどは放心状態で何も手につかなかったのですが、ポジティブに発想を転換させて、「フィトクロムの働きで転写開始点が変化するという現象が、GLYK遺伝子に限らず、それ以外の多くの遺伝子でも同様に見られたら、それはとても重要な発見になるに違いない」と自らに言い聞かせて、その可能性に賭けてみることにしました。当時、GLYK以外の研究はほぼ行っていなかったので、藁にもすがる思いがありました。
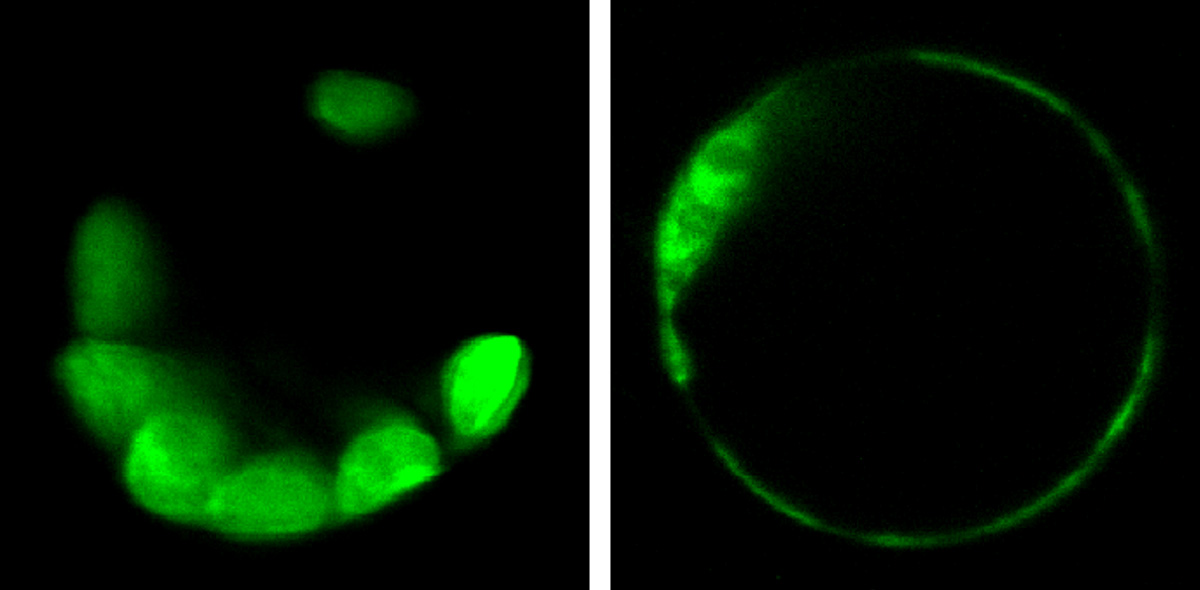
光依存的な転写開始点変化の発見
幸運なことにその仮説は当たっていました。再び花田さんの協力を得て解析を行ったところ、フィトクロムが転写量制御と選択的スプライシング制御に加えて、さらに転写開始点をも同様の規模で制御し、その結果数百という数のタンパク質の細胞内局在が赤色光依存的に変化していたのです。
さらにゲノム規模の転写開始点変化は、フィトクロムにより仲介される光刺激に限らず、ありとあらゆる環境刺激や信号に応じて起こりうるものであり、またその分子機構は真核生物に共通のものである可能性が高いことも分かってきました。つまり、フィトクロムの研究から偶然発見した転写開始点選択という現象は、実は真核生物のセントラルドグマにおいて、転写・スプライシング・翻訳と並んで第4の過程として働く普遍的な現象だったのです。植物のフィトクロムをひたすらズームアップしていった先に、忽然と普遍的現象の小宇宙が現れる・・・。それはまさに、子供の頃から植物に対して抱いていた浪漫が具現化された瞬間でした。

植物成長を支配する光
現在我々は、植物を題材にしてフィトクロムによる転写開始点制御の分子機構を解明することで、真核生物に共通と考えられる転写開始点選択のしくみを明らかにしたいと考えています。フィトクロムの研究をしていると、その制御の圧倒的な強さとそれの及ぶ範囲の広さから、植物にとって光というものがいかに重要なものであるのかを思い知らされます。今後も、植物における環境応答の1つの究極のモデルケースとして、フィトクロムを介した光応答の系にこだわり、マニアックに研究を狭く深く続けることで、その中に普遍的な現象の広がりをさらに見出していきたいと思っています。
構成協力/撮影 松林 嘉克
松下 智直 プロフィール:
1998年 京都大学農学部卒業。京都大学大学院理学研究科にて博士(理学)の学位を取得。米国UCLA研究員、九州大学農学研究院准教授を経て、2020年より現職。